Küresel Gıda Güvenliği İçin Genetik Mühendisliği: Fotosentez ve Biyofortifikasyon
Son 70 yılda, tarımsal alanlar küresel talebi karşılayacak düzeyde artmıştır. Bu artışların çoğu, tarımsal yaklaşımlardaki yüksek verimli çeşitlerin ve ilerlemelerin artmasından, ayrıca anahtar ürünlerdeki doğal fotosentetik varyasyondan yararlanarak verimi arttırmaktan kaynaklanmıştır. Fakat son günlerde bazı önemli gıda mahsullerinin verimleri durağanlaşmıştır.
2050 yılına gelindiğinde, küresel nüfusun 9 milyarı aşması beklenmektedir. Hem gıda hem de yakıt için sürekli büyüyen gereksinimleri karşılamak için, daha yüksek verime sahip yeni mahsul çeşitlerinin geliştirilmesi gerekli olacaktır. Bu ihtiyaçları karşılamak için buğday, soya fasulyesi (Glycine max) ve mısır (Zea mays) gibi mahsullerin veriminde % 70 ile % 100 arasında bir artış gereklidir. Mahsul üretimi için toprağı temizlemek, biyolojik çeşitlilikte küresel bir kayba neden olacak, böylece eşsiz habitatlara çevresel zarar dezevantajına sahip olacaktır. Bitkisel üretim için toprağı temizleyen bir yol, eşsiz habitatlarda çevresel hasara yol açmanın önemli bir dezavantajına sahiptir ve bu da biyolojik çeşitlilikte küresel bir kayba neden olmaktadır . Artan ekilebilir arazi, hem deniz hem de tatlı su ekosistemlerine daha fazla zarar veren ve sera gazı emisyonlarında önemli bir artışa yol açan gübre ve böcek ilacı kullanımının artmasına neden olmaktadır.
Çevresel zararı azaltmak için, ekilebilir arazi miktarını artırmadan küresel gıda taleplerini karşılamak gerekecektir. Çevreyi korurken verimi arttırmak için, yeni ana ürün çeşitlerinin geliştirilmesi esastır. Bu, genetik manipülasyon yoluyla fotosentezin iyileştirilmesiyle başarılabilir (Bölüm 2). Artan ürün verimine ek olarak, beslenme kalitesini iyileştirmeye de ihtiyaç vardır.
Son 5 yılda, yeni mahsul çeşitleri kalorili yoğun lifli mahsullerinin üretimini artırmıştır, ancak lif içermeyen mikro besin açısından zengin mahsullerin üretimi artmamıştır. Buna ek olarak, meyve ve sebze gibi lif içermeyen mikrobesin bakımından zengin ve önemli hayvansal ürünlerin fiyatları artmış, daha yoksul toplulukların sağlıklı bir diyet için gereken değişkenliği elde edememelerine neden olmuştur. Küresel olarak mikrobesin eksiklikleri iki milyardan fazla insanı etkilemektedir. Mikrobesin eksiklikleri, vitamin ve minerallerin sağlıklı bir yaşam sürdürmek için eşik değerin altında olduğu zayıf diyetin doğrudan sonucudur. Biyogüçlendirme sayesinde (Bölüm 3), yeni mahsullerde önemli besin maddelerinin biyoyararlanımını geliştirmek için potansiyel vardır.
Bu derlemede, fotosentetik verimliliğin artırılmasına yönelik çok hedefli bir yaklaşım ve çoklu vitamin bitkilerinin üretilmesine yönelik hedefli bir yaklaşım ile fotosentezin pozitif etkisinin olduğuna dair iddiaları ve modifiye edilmiş ürünlerin nihai vitamin içeriği üzerindeki etkisi arasındaki potansiyel bağlantıları tartışacağım.
FOTOSENTEZ
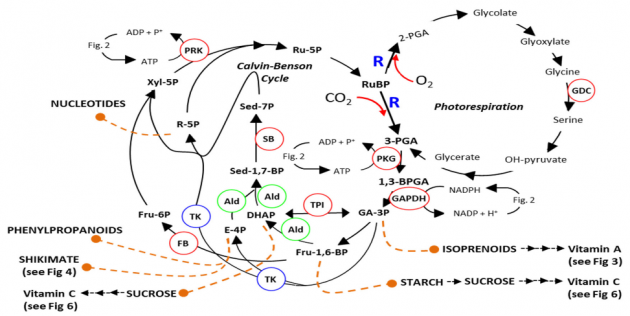
Fotosentez, dünya genelinde tüketilen tüm gıdalar için ya doğrudan bitki büyümesi yada dolaylı olarak gıda zincirinden sorumludur. Ayrıca, petrolden elde edilen tüm yakıt orijinal olarak fotosentlerden elde edilmiştir ve mevcut enerji üretiminin % 90’ını oluşturmaktadır. Mahsul verimi, büyüme mevsimi boyunca toplu fotosentez oranı ile belirlenir. Bunun önemli bir belirtisi, mahsül de A ışığının B’ye dönüştürülmesidir. (biyokütle veya tahıl) Bitkilerdeki karbon asimilasyonu, 1. Calvin-Benson çevrimi (CBC) ve 2. Fotorespiration olmak üzere birbiriyle yakından ilişkili iki yol içerir. Bu yollar, tüm yaprak proteininin % 50’sinden ve yaprak azotunun % 25’inden fazlasını oluşturan karmaşık bir stromal enzim olan Rubisco’nun (ribuloz-1,5-bisfosfat karboksilaz / oksijenaz; R; Şekil 1) katalitik aktivitesine odaklanmıştır. Tüm fotosentetik organizmalarda bulunan Rubisco, genetik manipülasyon için bir hedef olarak tanımlanmıştır ve bu nedenle bugüne kadar geniş bir şekilde ele alınmıştır.
Fotosentezi artıran elektron taşınması ve foto-solunum, mahsul verimliliğini artırmak için anahtar bir hedef olarak kabul edilmiştir. Simkin ve ark. [23] bu potansiyele değindi. Bu derlemede, dönüştürülmüş ürünlerin besin değeri üzerindeki potansiyel etkileri ve fotosentezi iyileştirmek için çok hedefli bir yaklaşımın etkisine odaklanacağız.
Calvin-Benson Döngüsünün (CBC) Manipülasyonu
CBC üç aşamaya ayrılır: karboksilasyon, indirgeme ve rejenerasyon. Melvin Calvin ve ark. 1950’lerde, 13 farklı adımı katalize eden 11 enzimi içeren döngüyü açıkladı. (Şekil 1). Transgenik çalışmalar, karbon asimilasyonu ve bitki büyümesi üzerinde önemli olumsuz bir etkiye sahip olan CBC’nin anahtar enzimlerinin hiçbirinde bir azalma göstermemiştir.
Bu çalışmalar sedoheptuloz-1,7-bisfosfataz (SBPase), fruktoz-1,6-bisfosfat aldolaz (FBPA), fruktoz-1,6-bisfosfatazlarda (FBPA) ( FBPase) veya fosforibulokinaz daki indirgenme nihai biyokütle veriminde bir azalmayla sonuçlandı. Bu deneylerin sonuçları, CBC içinde çok sayıda sınırlayıcı adım olduğunu ve bu dallanma noktalarının herhangi birinde artan enzim aktivitelerinin fotosentetik verimliliği artırma ve sonuç olarak verimi artırma potansiyeline sahip olduğunu göstermiştir. Ayrıca, saha bazlı çalışmalarda [CO2] zenginleştirme deneyleri bitki biyokütlesin de artışa neden olan artan CO2 asimilasyonunu göstermiştir.
CBC’deki tek adımları hedefleyen ilk çalışmalar, bunların bitki gelişimi ve biyokütle verimi üzerindeki etkilerini değerlendirme ve CO2 asimilasyon oranlarını artırmadaki potansiyelini test etmekte kullanıldı. Örneğin, tütün (Nicotiana tabacum), Arabidopsis (Arabidopsis thaliana) ve domatesdeki (Solanum lycopersicum) SBPase’in aşırı ekspresyonu, bitki biyokütlesinde önemli bir artışa (biyokütleden>% 30 artış) sebep olmuştur. Takip eden çalışmalarda, açık hava yükselen [CO2] deneylerinde yetiştirilen tütünün artan SBPase aktivitesi, biyokütle veriminde önemli artışlara neden olmuştur. Daha yakın zamanlarda, önemli bir gıda mahsulü olan buğdaydaki SBPazın aşırı ekspresyonu, biyokütle veriminde bir artışa ve daha da önemlisi, tohum veriminde% 30 ila% 40’lık bir artışa neden olmuştur. Benzer sonuçlar başka bir CBC enzimi FBPA’nın aşırı ekspresyonu ile elde edilmiştir, burada biyokütle sayısında% 70-120 oranında bir artış [CO2] (700 ppm) altında gözlenmiştir. Bununla birlikte, tütünle ilgili son çalışmalar, bazı CBC enzimlerinin aşırı ifadesinin yararlı olmadığını göstermiştir. Örneğin, transketolazın aşırı ekspresyonu bitki gelişimi üzerinde olumsuz bir etkiye yol açtı ve bitkiler düzenli olarak klorotik bir fenotip ve yavaş büyüme gösterdi.
Foto-Solunumun artışı verimi artırabilir mi?
CB döngüsünün bir parçası olan R ve kompleksi alt ürünlerdeki CO2’nin dahil edilmesi ile taşınan karboksilasyon reaksiyonları, CO2 yerinde R’nin O2 ile birleşmesinin olduğu yerde paralel bir rekabetçi reaksiyonla çelişmektedir (Şekil 1). Fotorespirasyon olarak bilinen bu reaksiyon, R tarafından gerçekleştirilen reaksiyonların yaklaşık% 25’ini oluşturur ve 3-fosfogliserat (3PGA) ve 2-fosfoglikolat (2PG) molekülünün oluşumuna yol açar. 2PG’nin oluşumu ve geri dönüşümü, fotosentetik verimliliği ve dolayısıyla verim potansiyelini olumsuz etkiler. Daha önceki bazı çalışmalar, yüksek fotorespiratuar koşullar altında (yani, yüksek sıcaklık veya su stresi) fotorespiratuar akıda bir azalmanın, fotosentetik verimlilik ve CO2 asimilasyonunda genel bir azalmaya neden olduğunu göstermiştir. Bunun aksine, glisin dekarboksilaz (GDC) H-proteininin veya L-proteininin yeşil dokuya özgü aşırı ekspresyonunun Arabidopsis ve tütünde bitkisel biyokütleyi arttırdığı gösterilmiştir. Fotorespirasyonun artmasının toksik fotorespiratuar ara maddelerin (örn., 2-PGA, glikolat, glikoksilat veya glisin) birikmesini önlediğine inanılmaktadır. Örneğin, 2-PGA’nın, bezelye [tr], trioz-fosfat izomerazı (TPI), ıspanakta fosforibulokinaz (PRK) ve Arabidopsis’te [70] SBPaz’ı inhibe ettiği gösterilmiştir. Ayrıca, Glyoxylate, ribuloz-bifosfat karboksilaz / oksijenaz (R) aktivasyonunu inhibe eder. GDC-H’nin aşırı ekspresyonunun son zamanlarda Arabidopsis’te fotosentetik elektron taşıma oranlarını arttırdığı gösterilmiştir.
Fotosentetik Elektron Taşımasının Geliştirilmesi Verimi Artırır
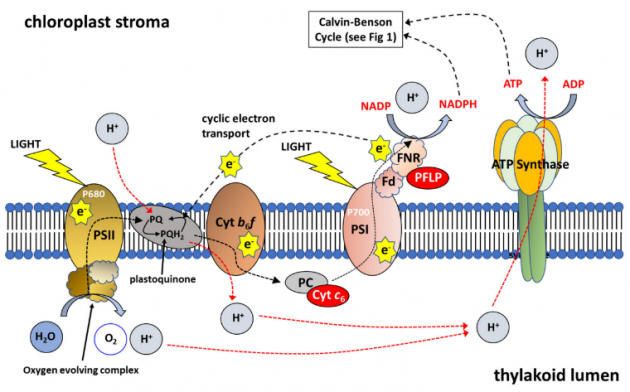
Fotosentetik elektron taşıma zinciri (ETC), ATP sentezi için gerekli bir elektrokimyasal proton gradyanı oluşturan fotosistem I (PSI) ve fotosistem II (PSII) arasında elektronların hareket ettirilmesi olan ardışık bir komplekstir. Alg (Porphyra yezoensis) sitokrom c6’nın ekspresyonu ile doğrudan artan fotosentetik elektron taşınmasının Arabidopsis’deki verimi arttırdığı gösterilmiştir(Şekil 2). Sitokrom c6’nın, bakır eksikliğine yanıt olarak siyanobakterilerde ve yeşil alglerde bir elektron taşıyıcı olarak plastokiyaninin (PC) yerini aldığı gösterilmiştir. Ayrıca Arabidopsis’te, cyt c6’nın sitokrom (cyt) b6f kompleksinden elektronları in vivo ve yerel PC için bildirilen oranlardan daha hızlı PSI’ya aktardığı gösterilmiştir.
Fotosentetik Verimliliği Artırmak için Çok Hedefli Yaklaşımlar Sinerjik Bir Etkiye Sahip olabilir
Yakın zamanda, CBC proteinlerinin, fotorespiratuar proteinlerin ve fotosentetik elektron taşınmasını arttıran proteinlerin aşırı ekspresyonunu birleştiren çalışmalar yapılmıştır. Bu sonuçlar çoklu hedeflerin manipüle edilmesinin biyokütle birikimi üzerinde sinerjistik bir etkiye sahip olabileceğini ve bu sinerjistik etkinin hedefe ve bitkiye özgü olduğunu göstermektedir (Tablo 1). Bu veriler ayrıca farklı yolları hedeflemenin de araştırılması gerektiğini göstermektedir. CBC’nin çift hedeflenmesi ve fotorespirasyon veya CBC ve elektron taşınması, hem kontrollü hem de saha deneylerinde verimde önemli artışlara neden olmuştur (bakınız Tablo 1). Bununla birlikte, ETC’nin hedeflenmesi şu anda tek hedef manipülasyonuna odaklanmaktadır (RieskeFeS, cyt c6 veya ferrodoxin: Şekil 2). Cyt b6f kompleksini, plaztosiyanin ve ferrodoksini aynı anda hedefleyerek elektron taşınmasını arttırmaya yönelik bir yaklaşımın, bir dizi çevresel koşulda elektron taşınmasını arttırma veya tutarlı yüksek oranları koruma potansiyeli de vardır. CBC, ETC ve Fotorespiration’u aynı anda hedeflemek, bitkilerdeki CO2 asimilasyon oranlarını iyileştirmek için bir sonraki mantıklı adımdır.
Fotosentetik Karbon Kullanılabilirliğinin Artırılması Vitamin İçeriğini (Biyolojik Güçlendirme) Olumlu Etkileyebilir
Son zamanlarda, yükseltilmiş [CO2] ‘nin ekşi portakal (Citrus aurantium) [98], çilek (Fragaria × ananassa) [99], domates ve havuçda ki (Daucus carota), C Vitamini miktarında (bkz. Bölüm 3.4) olumlu bir etkiye sahip olduğu bildirilmiştir. Elde edilen kanıtlara göre genetik manipülasyon yoluyla CO2 asimilasyon oranlarının arttırılması veya yüksek [CO2] ortamlarında (son günlerde domates ve çilek için kullanılan) yetiştirilen ürünlerin C Vitamini içeriğini potansiyel olarak etkilenebilir. (bkz. Bölüm 3.4) . Ayrıca, bitki dokularında C vitamini içeriğini artırma girişimlerinin ek sonuçları olmuştur. Vitamin C fotosentetik performansı ve büyümeyi arttırdığı gösterilen yeşil dokudaki c vitamini içeriğinin artmasında ve foto-korumada rol oynadığı gösterilmiştir.
Biyoforifikasyon
Biyoforifikasyon, yetiştirme veya genetik mühendisliği sayesinde bitki ürünlerindeki mevcut mikrobesinlerin arttırılması ve konsantre edilmesi işlemidir. Biyolojik olarak zenginleştirilmiş ürünler insan beslenmesini iyileştirmek için kullanılabilir ve beslenme değişikliği düşük uzak topluluklarda gerekli mikro besinleri sağlama potansiyeline sahiptir.
Mevcut yaklaşımlar büyük popülasyonların mikrobesin gereksinimlerini suni olarak karşılamak için hem işlendikten sonra gıdaya ekleyerek hemde hap şeklinde sunarak önemli ve sürekli bir finansal çıktıyı temsil etmektedir. Üretim veya genetik mühendisliği ile büyük popülasyonlara uzun bir süre boyunca besin takviyesi sağlama gereksinimi azaltılabilir. Çiftçilere ek bir maliyet olmadan besin açısından iyileştirilmiş ürünler yetiştirilebilir ve ilk araştırma yatırımı potansiyel ekonomik kazanımlara kıyasla küçüktür. Biyolojik olarak zenginleştirilmiş ürünlerin geliştirilmesine yatırılan her doların, 17 dolara kadar mali tasarruf sağlayabileceği tahmin edilmektedir.
Planta’da Pro-Vitamin A İçeriğinin Artırılması
A vitamini, en az bir β-iyonon halkası içeren C40 tetraterpenoidlerden (izoprenoidler; Şekil 1) elde edilir (bakınız Şekil 3A). β-karoten, β-kriptoksantin ve a-karoten yeşil sebzelerde ve meyvelerde bulunur. Bu üç karotenoid, insan diyetinde en yaygın A vitamini öncülerini temsil eder ve β-karoten, bitkilerde önemli bir foto-koruyucu olarak doğadaki en bol karotenoidlerden biridir. β-karoten, iki A vitamini molekülünün oluşturabilir ve tüm yeşil yapraklı bitki dokularında (yani marul (Lactuca sativa)) ve bazı meyvelerde (kayısı (Prunus armeniaca) ve şeftali (P. persica)) ve sebzelerde bulunur.
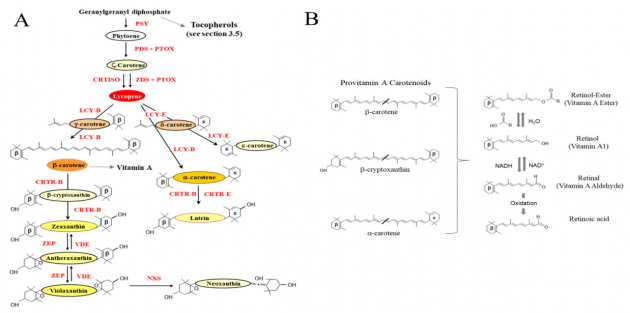
Karotenoid biyosentezinin genetik mühendisliği yoluyla bu bitkilerin A vitamini içeriğinin arttırılması, çeşitli domates (Solanum lycopersicum), mısır (Zea mays), buğday, kanola (Brassica napus), patates (Solanum tuberosum), keten tohumu (Linum usitatissimum), manyok (Manihot esculenta) ve Sorgum (Tablo 2)’daki yüksek karotenoid ile sonuçlanmıştır.
Oryza sativ (Os); Solanum lycopersicum (Sl); Capsicum annum (Ca); Brassica oleracea (Bo); Arabidopsis thaliana (At); Zea mays (Zm); Narcissus pseudonarcissus (Np); Hordeum vulgare (Hv); Musa troglodytarum x acuminate (Mt). CN = control.
Folat (B9 Vitamini) Birikiminin Genetik Manipülasyonu
Folat, bitkiler ve mikroorganizmalar tarafından sentezlenir, suda çözünür ve esansiyel bir mikro besindir. Folatlar bir pteridin, p Aminobenzoat (PABA) ve glutamat olmak üzere üç kısımdan oluşur (Şekil 4A). Bitkilerde, folat biyosentezi bölümlere ayrılmıştır, plastidde PABA (korisattan [Shikimate yolu, Şekil 1] ‘den türetilmiştir) ve sitozolde guanosin-50-trifosfattan (GTP)’den pteridinler sentezlenmiştir.
Bu iki kısım mitokondri içine taşınır, burada dihidropteroat oluşturmak üzere yoğuşturulur, bunlar daha sonra glutamatlanır (glutamat (Glu) ile reaksiyonla modifiye edilir) ve folatlar oluşturur (Şekil 4B). Bitkilerde, folatlar fotorespirasyonda ve klorofil, plastokinon, tokoferol sentezinde temel rol oynar, bu da onları bitki sağlığı ve gelişimi için gerekli kılar.
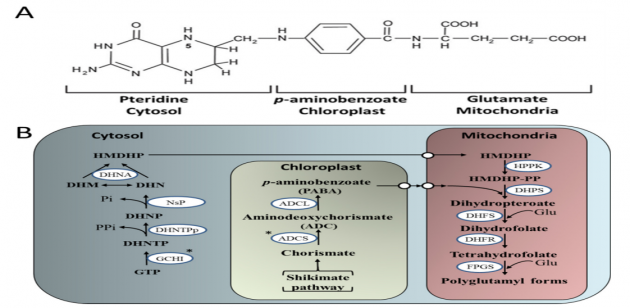
Pteridin yolunda [167] ilk önerilen adım olan GTP siklohidrolaz I’in (GCHI) (Şekil 4B) aşırı ifadesi (OE), Arabidopsis’te pterinlerde > 1000 kat artış ve sadece 2-4 – kat folatlarda artışla sonuçlanmıştır (Tablo 3). Ortak Meksika fasulyesinde GCHI’nın tohuma spesifik aşırı ekspresyonunun ardından benzer sonuçlar da gözlenmiştir [168]. Bu sonuçlar, domates meyvesindeki GCHI OE’sinin pterinlerde 140 kata kadar artış ve meyve folat içeriğinin ortalama 2 kat arttığını gösteren Diaz de la Gaza tarafından yapılan benzer çalışmalarla desteklenmiştir. Domates meyvesinde, GCHI’nin OE’sinin, transgenik domates meyvesinde folat biyosentezinde bir başka sınırlayıcı aşamayı temsil ettiği kloroplast sentezlenmiş PABA’nın tükenmesiyle sonuçlandığı gösterilmiştir (Tablo 3). Bununla birlikte, PABA’daki bu tükenme Meksika fasulyesinde GCHI’nin OE’sini takiben gözlenen bir artışla zıttır. Bu yazarlar meyve sapı olsa da meyve PABA’nın beslenmesinin folatı 10 kata kadar artırabildiğini göstermişlerdir.
Meyve folat içeriğini arttırmak için başka bir girişim, plastid lokalize aminodeoksikrizmat sentazın (ADCS) (Şekil 4B) OE’sindedir ve olgun meyvede PABA seviyelerinde 19 kata kadar artış sağlanmıştır. Diğer türlerde yapılan çalışmalar, metabolik mühendislikle artan folat konsantrasyonunun, gelişmekte olan ülkelerde tüketilen temel ürünler de dahil olmak üzere daha sağlıklı takviye edilmiş ürünler üretmek için önemli bir yol olduğunu göstermiştir (Tablo 3).
Kobalamin (B12 Vitamini)
Biyolojik olarak aktif B12 Vitamini (B12) formu olan kobalamin, klorofil ve hem haem gibi molekülleri içeren modifiye edilmiş tetrapyrol ailesinin bir parçasıdır ve sadece küçük bir grup prokaryot (bakteri ve arkea) tarafından yapılır. Bu bakterilerin bazıları, midede çoğaldıkları ve B12 oluşturmaya devam ettikleri geviş getiren memelilerin florasında bulunur. Bu B12 kaynağı et, yumurta, süt dahil olmak üzere hayvansal üründe birikir ve popülasyondaki B12’nin temel diyet kaynağıdır ve bu da çoğunlukla takviye edilmiş tahıllar, bitki ve besin takviyesi gerektiren vegan diyetini takip edenler için sorunları gündeme getirir. ciddi sağlık sonuçlarından kaçınmak için süt sağımları, beslenme mayası, tabletler veya kas içi enjeksiyon. B12 meyve ve sebzelerde bulunmaz, ancak yenilebilir mor / yeşil nori önemli miktarda B12 içerir. Bununla birlikte, bu B12 kaynağı, biyoyararlanamayan ve aynı zamanda insan diyetlerinde aktif olmayan bir analog olan yüksek miktarlarda psödovitamin B12’nin varlığı nedeniyle tartışmalıdır.
Yetişkin erkekler ve kadınlar için önerilen günlük B12 ödeneği, hamile kadınlar için 2,6 µg / gün ile 2,0–2,4 ug / gündür. B12 vitamini, DNA ve nörotransmiterlerin yanı sıra yağ asidi ve amino asit metabolizması için gerekli bir kofaktör olduğu hayvanlar için gerekli bir besindir. Normalden biraz daha düşük seviyelerde bile, B12 eksikliği depresyon, hafıza kaybı (düşük bilişsel performans), yorgunluk, uyuşukluk ve baş ağrıları ve bazı insanlarda mani ve psikoz gibi bir dizi semptomla sonuçlanabilir. Bu semptomların çoğu, pernisiyöz anemi, göğüs ağrısı, el ve ayaklarda uyuşma, zayıf refleksler, pürüzsüz kırmızı dil ve nefes darlığı gibi ek semptomlara yol açabilen kırmızı kan hücrelerinin eksikliğinin sonucudur.
Kobalamin, merkezinde dört pirol halkası (Şekil 5) ile çevrili bir kobalt içeren en yapısal olarak karmaşık ve en büyük vitamindir ve biyosentezi yakl. Ökaryotik hücreler tarafından sentezini oldukça zorlaştıran bir dizi oksijene duyarlı ve oldukça kararsız ara madde dahil olmak üzere 30 enzimin aracılık ettiği aşama.
L-askorbik Asit (C Vitamini)
İnsanlar ve diğer bazı primatlar, C vitamini sentezleme ve depolama yeteneğini kaybettiler ve diyetlerinde günlük 75-90 mg’lık ihtiyaçlarını karşılamak zorundadırlar. C vitamini (Şekil 5) turunçgiller (portakal ve limon vb.), Greyfurt, mango, kivi, brokoli, Brüksel lahanası, domates ve çeşitli meyve meyvelerinde (çilek, ahududu, yaban mersini ve kızılcık) bulunur. C vitamini, bağ dokularının onarımı, kollajen sentezi ve bazı nörotransmitterlerin enzimatik üretimi için gerekli olan temel bir besin olarak kabul edilir. Bağışıklık sistemi fonksiyonunun doğru çalışması için önemlidir ve bir antioksidan olarak görev yapar. Önceki araştırmalar gösterdiki, C vitamininin Alzheimer, Huntington ve Parkinson hastalığının tedavisinde yardımcı olabilir.
Fizyolojik pH koşulları altında, C vitamini özellikle metilsitozin dioksijenazlar için bir kofaktör olarak, DNA’nın demetilasyonunda yer alan enzimler için bir askorbat anyonu ve fonsiyonlar olarak işlev görür. Ayrıca, Askorbat, histon demetilazyonu için gerekli Jumonji C-domain içerikli histon demetilazları için bir kofaktör görevi görür. Bu bilim adamları, C vitamininin, çevre ve genom arasındaki etkileşime aracılık eden epigenetik süreçleri düzenlediğini hem DNA’nın hem de histonların demetilasyonuna katıldığını öne sürdüler.
Bitkilerde, Askorbat metabolizması fotosentez ile güçlü bir şekilde bağlantılıdır ve fotokorumada rolü ispatlanmıştır. Askorbat, kloroplastta önemli bir antioksidandır ve karotenoid biyosentetik yolağındaki VDE aktivitesi için önemli bir kofaktör olduğu bildirilmiştir (Şekil 3A). Işığın soğurulması bitkinin kapasitesinin ötesinde olduğu zaman, fazla enerjinin dağıtılması gerekir.
Bitkilerde, birkaç farklı askorbat biyosentez yolu karakterize edilmiştir (Şekil 6). Smirnoff-Wheeler yolu bitkilerdeki birincil yoldur, ancak üç ‘alternatif’ yolun da plantada askorbat üretimine yol açtığı gösterilmiştir. Miyo-inositol yolu, L-guloz yolu ve D-galakturonat yolu.
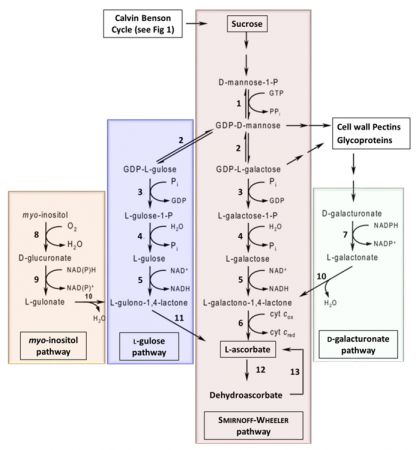
Hem Smirnoff-Wheeler hem de L-gulose yollarında önemli bir adımı olan GDP-Man-3 ‘, 5’-epimeraz ((GME) enzim 2: Şekil 6 ve Tablo 4) genlerinin aşırı ekspresyonunun, domates meyvesinde ki askorbatta 1.2 den 1.6 kata kadar artışla sonuçlandığını gösterir.
Ayrıca, hem Smirnoff-Wheeler hem de L-gulose yollarının her ikisinde de aktif olan aşırı ekspresyonlu GDP-L-Gal fosforilaz ((GMP) enzimi 3: Şekil 6 ve Tablo 4), çilek ve domates meyvesinde askorbat seviyelerinde 2- ile 6 kat artış ve patates yumrularında askorbatta 3 kat artış ile sonuçlanmıştır.
GDP-L-Gal fosforilaz ((GGP) enzimi 3: Şekil 6) aşırı ekspresyonu askorbat içeriğini arttırdı, bundan dolayı GGP’nin Smirnoff-Wheeler yolunda önemli bir enzim olduğunu düşünülmektedir. Diğer çalışmalar GDP-D-Man pirofosforilaz ((GMP) enzimi 1: Şekil 6) ‘nın aşırı ekspresyonunun da askorbatta ki artışı kolaylaştırdığını göstermiştir.
Tokoferoller (E Vitamini)
E vitamini, en aktifi a-tokoferol olan dört tokoferol ve dört tokotrienol içeren yağda çözünen bir bileşiktir (Şekil 5). Hücre zarlarını, sinir hasarına yol açabilen reaktif oksijen türlerinden koruma işlevi görür. E Vitamini eksikliği insanlar nadirdir ve genellikle diyette E Vitamini eksikliğinden ziyade metabolik bir bozukluğun sonucudur. Mısır’daki E Vitamini içeriğini arttırmak için Cahoon ve ark. Homogentisik asit geranilgeranil transferazı (HGGT) kodlayan cDNA’ların aşırı ekspresyonu, toplam E vitamini antioksidanlarında (tokotrienoller artı tokoferoller) 10-15 kat artışa neden olmuştur.
Bunu karotenoid biyosentetik enzimleri kodlayan cDNA’nın aşırı ifadesi ile kombinasyon halinde HGGT’yi aşırı eksprese eden (Tablo 2) Sorghum’da Che ve ark. kanıtlamıştır. Bu yazarlar, bu gen kombinasyonu ile transgenik sorgumun all-trans β-karoten (~ 19 kat artış bkz. Bölüm 3.1) ve a-tokoferol, γ-tokoferolde sırasıyla 1.8 kat ve 1.7 kat ve a-tokotrienolde 27 katlık bir artış sağlayabildiğini göstermiştir.
Çoklu Vitamin Ürünlerine Çok Hedefli Bir Yaklaşım
Önceki çalışmalar, çeşitli ürünlerin besin içeriğini arttırmak için çoklu genlere hedeflenme yaklaşımını kullanmışlardır; bununla birlikte, Naqvi ve ark., Güney Afrika mısır’ında C vitamini, folat ve provitamin A olmak üzere üç farklı metabolik yoldan üç vitamini artırmak için multigen, multivitamin yaklaşımı kullandılar.
Gelecekteki Beklentiler ve Sonuçlar
Gıda ve yakıta olan artan gereksinimler ile daha fazla verim ile önemli mahsul çeşitlerinin geliştirilmesi ihtiyacı tarımın ön saflarında yer almaktadır. Son yıllarda, verimi arttırmanın bir yolu olarak fotosentez oranını arttırmak için genetik manipülasyon kullanılması da ivme kazandırmıştır. Bunu destekleyen kanıtlar, transgenik bitkilerin değişikliğinde tekli genlerin aşırı ekspresyonuna bakarak hem modelleme yaklaşımlarından hem de ilk çalışmadan ileri gelmiştir.
Daha sonra, genetik manipülasyona yönelik çok hedefli bir yaklaşım, CBC, fotorespirasyon ve elektron taşınmasında birden fazla hedefin manipüle edilmesinin, verimde daha fazla artışa neden olan sinerjik bir etkiye sahip olabileceği fikrini de destekledi. Ayrıca, artan [CO2] ‘de büyüyen havuç, çilek, domates, kereviz ve narenciye, C vitamini içeriğinde artışa neden olarak, artan CO2 asimilasyonunun beslenme içeriğini artırma potansiyeline sahip olduğunu gösterir. Bu veriler ayrıca atmosferik [CO2] ‘yi arttırmadan CO2 asimilasyonunu iyileştirmek için fotosentezin manipüle edilmesinin potansiyel olarak bitki vitamin içeriğinde de artışlara neden olabileceğini düşündürmektedir. Ancak, bu durum daha karmaşık ve türe özgü olabilir. Cober ve diğ., artan [CO2] ‘de soya fasulyesinin büyümesinin tohum veriminde bir artışla sonuçlandığını, ancak tohum yağı konsantrasyonlarının ve tohum proteini seviyelerinin azaldığını bildirdi. Ayrıca Myers ve ark. Bir meta analiz yaparak yüksek [CO2] altında büyütülen C3 bitkilerinin ve baklagillerin daha düşük çinko ve demir konsantrasyonlarına sahip olduğunu ve baklagil olmayan C3 bitkilerinde protein içeriğinin azaldığını bildirmişlerdir. Zhu ve diğ. bu sonuçları doğruladılar ve ayrıca yüksek [CO2] altında yetiştirilen Pirinç’teki B1, B2, B5 ve B9 vitaminlerinde düşüş olduğunu gösterdiler. İlginç bir şekilde, bu yazarlar aynı bitkilerde E vitamini artışı (bkz. Bölüm 3.5) gözlemlediler. Atmosferik [CO2], sıcaklık ve yağıştaki küresel artış iklim değişikliğini gösterir, bunun sonucu da ürünlerimizin beslenme kalitesini önemli ölçüde etkileyebilir.
Biyoteknoloji programları tarımsal araştırmalarının ön safhasıdır, örneğin genetik mühendisliği ve endojen genlerin manipülasyonu için genom düzenleme gibi tekniklerin benimsenmesi bu sorunların üstesinden gelmenin bir anahtarıdır.
Verim ve beslenme kalitesinde sürdürülebilir artışlara sahip ve değişen çevresel koşullarına başarılı bir şekilde adapte olabilen bitkiler üretmek için, karbon asimilasyonu, ışık adaptasyonu ve besin biyosentezinin birçok yönüne dokunan çok hedefli bir yaklaşım ve çoklu gen sokulmasında ihtiyaç duyulan vektörler ve dokuya özgü promoterler için yeni araçlar gerekli olacaktır.
Önceki çalışmalar, provitamin A, Folat ve C vitamini metabolizması için genlerin ekspresyon seviyelerinin manipüle edilmesini göstermiştir ki buda, bu mikrobesinlerde bir artışa ve multivitamin mısır da bu manipülasyonların birleştirilmesi ile sonuçlanır. Bu araştırma, risk altındaki popülasyonlarda “açlık” ve “gizli açlık” konusuna yönelik yüksek verimli multivitamin mahsulleri oluşturmak için fotosentez ve vitamin metabolizmasının manipüle edilmesi yolunu açar.
Çeviri Kaynak: https://doi.org/10.3390/plants8120586